This has to be quick, as I'm close to emerging from seemingly never-ending book revision and want to finish it as soon as possible, but something of interest to Pterosaur.Net readers (and, indeed, anyone with an interest in palaeontology, natural history or science communication) has emerged that needs bringing to the widest possible attention. Over at Tetrapod Zoology, Darren Naish has recently published a long, detailed
critique of the many problems inherent with a website with worrying Internet presence, David Peters'
ReptileEvolution.com. The title of the piece, "
Why the world has to ignore ReptileEvolution.com" probably tells you everything you need to know about its content but, if that doesn't spell it clearly enough, its loud message at the top of the page makes the point clearer:
In all likelihood, if you're reading this post, you are already familiar with Peters, his website and its sister blog,
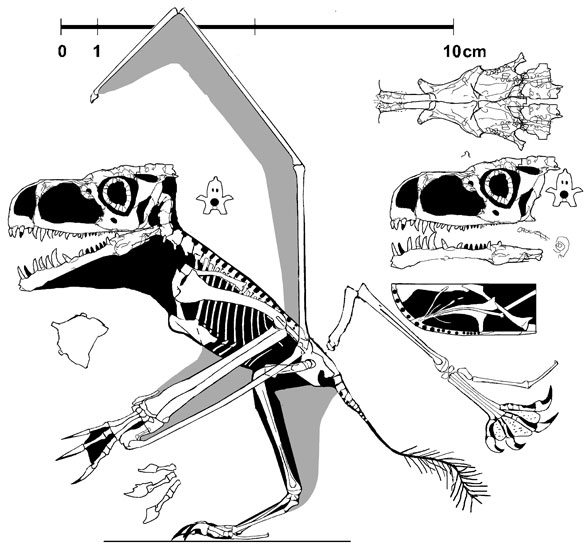
The Pterosaur Heresies. Even if these names are not familiar however, there is a good chance you have bumped into them when Googling almost any Mesozoic reptile you care to think of. Peters is well-known in palaeontological circles (though perhaps mostly ignored by the professional palaeontological community now) for his unorthodox views on amniote phylogeny and, perhaps more commonly, his sometimes bizarre interpretations of pterosaur anatomy and functional morphology. For years, Peters has been using a technique known as "Digital Graphic Segregation", tracing photographs of fossils and interprets actual bone, marks in the surrounding matrix, and probably preparation, printing and jpeg compression artifacts, to reconstruct the anatomy of fossil animals. Observations on actual specimens are very much of secondary concern and do not factor into this technique much, if at all. This leads to the frequent identification of features that simply do not exist on the actual specimens (see Peters' reconstruction, above, of
Anuroganthus ammoni for an example [
source]
. Note the number of phalanges in the wing finger, shape of the skull and long, fibrous tail. Go
here to compare this with the best-preserved specimen of this pterosaur) and, when fed into a phylogenetic analysis, the resultant trees are understandably completely incongruous with anything seen in 'mainstream' literature. Peters' work has been rightly criticised from all angles (including Bennett 2005; Hone
et al. 2009,
this, and frequently on the Dinosaur Mailing List) but he retains his ideas in the face of overwhelming evidence to the contrary (i.e. the inability to see the structures he claims to find on actual specimens despite microscopic, and UV observation, and CT scanning). As such, the work portrayed on his website and blog has to be pseudoscience, at best.
Above, ReptileEvolution.com image allegedly demonstrating the presence of a fifth wing phalanx in the pterosaur wing. Consistent observations of pterosaur fossils, by contrast, suggest they only had four phalanges, or sometimes three. From here.
This, in itself, is not really the problem. As Darren points out, the Internet is a place for creative, free thinking individuals and, hey, if you cannot express your opinions here, where can you? The issue taken with ReptileEvolution.com is not that it exists, but that it's internet presence has grown to the point that it is now a top-listed site for many palaeo-based searches. Tap virtually any Mesozoic reptile species into Google and either ReptileEvolution.com or the Pterosaur Heresies is likely to be in the first few hits. The situation is even worse for image searches, which are increasingly dominated by the many graphics that Peters' uses on his sites. This would seemingly be because Peters is extremely prolific in his output on these projects, and because most Mesozoic reptiles are poorly covered online.
Those In The Know are, at worst, merely frustrated by this situation, as it skews search results from other hits that may be of interest. Those who are not familiar with Peters' work or sites however, are potentially being mislead by his very professional-looking websites that frequently present their content as hard biological facts rather than the eccentric views of one individual. I have heard anecdotes of news outlets citing ReptileEvolution.com and, not many months ago, we ran into a
direct example of one website blindly following Peters' ideas (
we weren't impressed). This is a worrying trend.
We have more-or-less ignored Peters' work at Pterosaur.Net, which may seem surprising given that he has dedicated a whole blog, more or less, to deconstructing the 'mainstream' view of pterosaur palaeobiology. ReptileEvolution.com even has a page dedicated to
picking holes in the main Pterosaur.Net site! I suppose we have more interest in discussing other topics on our blog than merely rehashing the same anti-Peters arguments over and over. Most of us have had discussions with Peters about his ideas and hypotheses at one venue or another anyway, so I guess we've 'moved on' in some respects. Regardless, I'm sure I speak for my colleagues here at Pterosaur.Net when I say that we are as concerned over the prevalence of ReptileEvolution.com and the Pterosaur Heresies as Darren is at TetZoo, and want to bring these concerns to the widest possible audience. Hence, I urge you to read Darren's
discourse if you have not already done so and, if you are concerned about the accurate portrayal of palaeontological science online, then blog, tweet and discuss this issue as much as you see fit. As may be expected, Peters has started a rebuttal of the piece across a number of blogposts, which begins
here.
That will have to do: I've gone on far longer than planned. Back to the book...
References
- Bennett, S. C. 2005. Pterosaur science or pterosaur fantasy? Prehistoric Times, 70, 21-23.
- Hone, D. W. E., Sullivan, C. and Bennett S. C. 2009. Interpreting the autopodia of tetrapods: interphalangeal lines hinge on too many assumptions. Historical Biology, 21, 67-77.